Distribution of the Phasmatodea
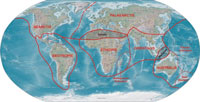
Zoologists, studying the distribution of animals (zoogeography or biogeography) divide the world into six well defined zoogeographical regions or faunal realms, in each of which at least 50 % of the known mammals are endemic (see BRIGGS, 1987, COX & MOORE, 1993). These are the Palaearctic, Nearctic, Australian (= Notogaea), Afrotropical, Oriental and Neotropical regions (► map 1). As the Nearctic and Palaearctic regions exhibit very similar and closely allied faunas they are often combined as the Holarctic region. This is a similar case with the Afrotropical and Oriental regions, which are often combined as the Palaeotropics. Major zones of transition whose faunas cannot definitely be assigned to a certain zoogeographical region are marked by hatching in the map below (► map 1). These are the Sahara and the Wallacea. The latter contains the Lesser Sunda Islands (Lombok, Sumbawa, Sumba, Flores, Wetar & Timor), the Philippines, Sulawesi (= Celebes), the Moluccas, Ambon, Ceram as well as the Key Island. Whether the Philippines should be included in the Wallacea is still subject of discussion (see VANE-WRIGHT, 1990, HUXLEY, 1868). The “Wallace Line” represents the western boundary of the Wallacea, but the eastern boundary is still not a confirmed but generally accepted to be Weber’s Line (see SIMPSON, 1997).
Madagascar, Seychelles, Comoros and the Mascarene Islands form a quite well-separated subregion of the Afrotropical region, which lacks most of the typical African faunal elements. Its fauna shows distinctly more close relations to that of the Oriental and Australian regions rather than to the African continent. Other well-defined sub-regions with rather distinctive faunas are the Caribbean subregion (part of Neotropical region), as well as New Zealand or the Papuan and Polynesian subregions of the Australian region (► map 1).
The English zoologist Alfred Russel WALLACE (1823-1913) was the first to define the division of the earth into zoogeographical regions based on the mammal fauna in 1876, based on the mammal fauna. Although WALLACE’s concept has seen numerous additions and sub-divisions into subregions or zoogeographical provinces during the past 140 years, his division has principally remained valid until now.
The Phasmatodea distribution areas encompass most parts of the warm climate zones of all zoogeographical regions, but especially the tropical and subtropical areas. The Oriental and Neotropical regions do by far represent the richest and most diverse phasmid faunas, followed by the Australian region, Madagascar, the Ethiopian, Nearctic and finally the Palearctic region (► map 2 below).

The habitats in which phasmids occur can be divided into groups due to their diverse vegetation and climatic conditions. The identification of these habitat types requires knowledge of the typical tree and plant species. These have a great influence on the spectrum of phasmid species occurring in a particular habitat. In addition, the different habitats show a close connection to the vertical height and vegetation zones and the associated climatic conditions. In the following, different habitats are briefly described and some habitat-typical phasmid species are mentioned.
Mangrove Forest
Mangroves are evergreen forests with only a few different tree species that settle in sheltered bays, lagoons and river mouths within the tidal range and prefer clayey sandy swamps. These areas are flooded at high tide, dry at low tide and are largely dominated by tree-like halophytes (salt-tolerant plants) with their typical stilt and breath roots. The largest mangrove diversity is reached in the Oriental Region. The phasmid fauna of the mangrove forests is sparse and mostly limited to a few species. For example, at "Cape Tribulation" in north-eastern Australia (Queensland) only Megacrania batesii (Kirby, 1896) occurs, which can be found on Pandanus palms (Pandanaceae, screw pine).
Riparian Forest / Floodplain Forest
Tropical floodplain forests are those lowland forests that are flooded at least once a year for more than two days. In most cases, these areas grow few trees and are characterized by low plant diversity. Since the water flows off more quickly in the immediate vicinity of rivers and therefore the flooding periods are shorter than in the other areas, the phasmid fauna along rivers is often not necessarily typical for the adjoining floodplain forest area. There are normally no species in these areas which lay their eggs into the soil or drop them to the ground, as these would be flooded during the flood periods. The phasmids found there are usually flying species which stick or prick their eggs to branches or leaves of the food plant or lay them under tree bark. The species density is generally quite low, but increases near rivers.
Tropical Lowland Rainforest
Tropical lowland rainforests cover large areas in Southeast Asia, North Australia and the Amazon lowlands of South America. They grow only in hot tropical climates, where the average annual temperature fluctuates only slightly (between 24 and 30°C) and even the coldest monthly average does not fall below 18°C. The temperature in the tropics is very low and the average temperature in the tropics is very low. Rainfall in these areas is high (2000-6000 mm per year) and often evenly distributed over the seasons. At certain times of the day, there are regular heavy downpours, and longer dry periods are usually completely absent. Tropical lowland rainforests can be found up to 1500 m above sea level. The constantly high humidity and the balanced temperatures allow lush vegetation to thrive, although the characteristic red clay soils have a low humus content and therefore few nutrients. The soils of the tropical rainforest zone are among the most infertile on earth.
The tropical rainforest is incredibly rich in evergreen trees but almost free of conifers. Botanists estimate that two-thirds of all plants on earth occur in the evergreen tropical rainforest. In the Amazon lowlands, for example, only a few hectares of forest are home to more tree species than the whole of Europe (60-200 species per hectare), and it is not unusual for a single tree to be populated by up to 100 different species of climbing plants and mosses. The multi-layered, vertical structure of lowland rainforest is very complex (see figure 2 below) and is divided into:
- shrub- and herb-layer
- understory layer
- canopy layer
- emergent layer
The canopy layer, which forms a dense canopy of leaves at heights of 20-30 m and allows only a little sunlight to penetrate to the rainforest floor, is only interrupted by giant trees up to 60 m high (emergent layer). The undergrowth (understory layer) consists mainly of young plants of the dominant trees and shrubs as well as various giant shrubs (e.g. Musaceae). The shrub and herb layer is mostly formed by ferns and other shade-loving plants with often colourful leaves. The "striving for light" is particularly expressed by the abundance of lianas, epiphytes and other climbing plants.
The lowland rainforest is home to a wide range of different phasmid species and groups. Since the lower, easily searchable vegetation layer is rather meagre, it is sometimes difficult to find phasmids in such areas. Most species are forced to look for their forage plant in the understory and canopy layers, where they are hardly accessible for phasmatologists without appropriate collection or climbing equipment. Although the shrub and herb layers also contain various flightless species, their numbers are small compared to secondary or mountain rainforests. The largest part of the phasmid fauna in these areas consists of small, well-flying species in the treetops (canopy layer). The most abundant collection sites in the primary lowland rainforest can be found along rivers, the edges of deforested areas or along roads, logging and hiking trails.
The upper parts of the rainforest (including the treetops) are still largely unexplored and are therefore an interesting area of research for many biologists. New insect species are discovered almost daily in the treetops of tropical rainforests in different parts of the world.

Phasmid species in the different zones:
- shrub- and herb-layer (0 - 5 m)
small to medium-sized, wingless and isolated, flying species as well as various larvae,, e.g, Dares, Pylaemenes, Hoploclonia, Aretaon, Haaniella, Acanthoclonia, Autolyca, Creoxylus, Parectatosoma, Eurycantha, Libethra, Oreophoetes, Medaura, Ramulus, Dyme, Calynda, Bostra, Lonchodes, Pseudophasma - understory layer (5 - 10 m)
larger, flying species and giant stick insects, e.g. Orthonecroscia, Calvisia, Necroscia, Diesbachia, Citrina, Pseudophasma, Paraphasma, Prexaspes, Bostra, Phanocles, Pharnacia, Phobaeticus - canopy layer (4 - 15 m)
various flying species and giant stick insects, e.g. Necroscia, Stratocles, Melophasma, Prisopus, Paraphasma, Pseudophasma, Pharnacia, Phobaeticus, Hermarchus, Ctenomorphodes, Acrophylla, Tropidoderus, Extatosoma, Cladomorphus - emerging layer (15 - 50+ m)
small to very small, flying species and changing leaves, e.g. Necroscia, Nescicroa, Stratocles, Phyllium
Vegetation:
A: soft-leaved herbs and perennials (e.g. Araceae: Begonia, Dieffenbachia & Spatiphyllum)
B: Fast-growing trees (e.g. Cecropia spp., Moraceae)
C: Young shrubs and trees
D: Climbers (e.g. Calamus spp.)
E: Ferns and tree ferns
F: Climbing plants (e.g. Araceae: Epipremnum, Scindapsus & Philodendron)
G: Giant tree (over 50 m) with board roots
H: Epiphytes (e.g. Bromeliaceae, Orchidaceae)
J: Palm
K: Staff trees with stilt roots (e.g. Ficus)
L: Upper tree layer
Tropical lowland secondary forest
Tropical lowland secondary forests and mountain rainforests are the most productive habitats when searching for phasmids. Usually, representatives from all important groups are present there. Secondary forests are formed by deforestation and slash-and-burn of primary forest areas, which are then used for agricultural purposes for several years. If such an area no longer yields enough crops to secure a livelihood, it is quickly abandoned and a new forest, the secondary forest, can slowly form. Usually, the trees are younger and considerably lower than in the surrounding primary forest. Therefore, considerably denser and richer vegetation can develop near the ground. In the immediate vicinity of the primary forest, such areas provide a very suitable new habitat opportunity for various animals, including phasmids. Secondary forests that have grown for at least 10 years are often very productive collection sites for phasmids. Just the access to secondary forests is often somewhat difficult, due to the dense low-level vegetation, yet there is plenty of vegetation to be searched. Additionally, near agricultural areas, there are often paths alongside and through the thicket, which make collecting much easier. Especially the edges of secondary forests have proven to be extremely successful and productive. Thickets along major roads, which cut broad forest aisles into the primary rainforest, are fauna-rich secondary forests too.
Since the vegetation grows lower than that in the primary forest, it is possible to find species here which otherwise occur mainly in the treetop layer. Typical trees of the secondary forest in South America are for example the fast-growing trees of the genus Cecropia (Moraceae).
Tropical Mountain Rainforest
Tropical mountain rainforests are found between 1500-2500 m above sea level. They are similar in structure to tropical lowland rainforests but are characterized by a lower tree layer (5 - 15 m) and denser shrub and herb layers. Tree ferns and epiphytic mosses emerge in large numbers. There are also various palms and conifers (conifers such as Podacarpus spp. in South America) as well as many orchids and bromeliads (in South America). Due to orographic rainfall, precipitation is even more frequent here than in the lowland rainforest. Due to the frequent precipitation, the mountain slopes favour landslides, which tear large aisles into the vegetation cover. This creates new habitats for many pioneer plants and subsequently diverse and dense vegetation. Mountain rainforests, therefore, have a very high biodiversity (species density). In addition, there is the condensation of fog droplets on branches and leaves, so that the vegetation is exposed to high humidity in a rather cool climate all year round. Besides secondary rainforests, tropical mountain rainforests are the most productive and easily accessible areas for phasmid collection. Because of the lush, low vegetation, the highest density of phasmid species can also be found here.
Could Forest
A special form of mountain rainforest is the tropical cloud forest. In principle, cloud forest is a sub-concept for different (altitude) levels of the mountain rainforest and is located between 1500 and 3000 meters. At this altitude, the cloud level is reached, and thus the zone of maximum humidity. Here the cloud forest spreads out, which depends on almost 100% humidity. The extreme humidity and mild temperatures in connection with sufficient sunlight allow for vegetation, which resembles the mountain rainforest but has fewer species. The dominant plants here are tree ferns, epiphytes as well as moss and liver moss carpets, which spread over the entire soil and also envelop the trees and shrubs in a green overgrowth. Often there are different begonias, fuchsias, orchids or bromeliads.
The phasmid fauna of the cloud forest is similar to that of the mountain rainforest but has fewer species. Collecting can be difficult due to the dense fog.
Dwarf Forest
Starting from heights of 2500 m, mostly above the cloud zone and above the tree line, one can find dwarf forests. With relatively little rainfall, compared to the regions at lower altitudes, the green of the trees becomes sparser and the maximum tree height is barely more than 5 metres. In the Andes of South America, the tree layer of the dwarf forest is composed almost exclusively of Podacarpus species (Podacarpaceae), while in Africa at this altitude mainly plants of the Ericaceae family can be found. The leaves of these shrubs are always hard and small. Mosses are replaced by beard lichens. The vegetation near the ground is dense and almost impenetrable, but not very diverse.
The phasmid fauna of the dwarf forest zone is generally quite poor, but especially in the dwarf forest zones of the Andes in Colombia and Ecuador a surprising number of often strongly adapted phasmid groups occur. Due to the low vegetation and the often isolated growing "forest islands", the collection in these areas is simple but, apart from certain regions of the Andes, not very productive.
Páramo (Alpine Tundra / Highland Steppe)
The tropical highland zone of the Páramo in the Andes of Latin America is unique in the world. This wet and cold grassland zone is found exclusively between 10° northern and 10° southern latitude, i.e. from the highlands of Costa Rica to the north of Peru, and at altitudes between 3000 and 4700 m. The extremely specialized Páramo vegetation has developed over the course of more than 60 million years and forms mostly heather-like formations of hedgehog grass (Stipa ichu, Gramineae), nosegrass, small laurel-leaved shrubs as well as low cushion and rose plants (e.g. Polylepis spp., Rosaceae). Typical for this region are the Espeletia spp. with a height of about 1-8 m and Puya spp.. The leaves of the Páramo plants are small, and thick and often arranged in symmetrical rosettes, which results in better absorption of sunlight. A thick wax layer protects the leaves against the intense sunlight and small hairs protect them against the cold and store the necessary warmth. The flat growth of most plants provides lesser exposure to wind and cold. The Páramos have an extreme daytime climate with frequent precipitation. A strong wind blows, which often increases to snow and hail storms and passing clouds can cause temperature drops of up to 15°C in only a few minutes. While temperatures can rise to over 20°C during the day due to intense sunshine, night frost can occur almost daily at altitudes above 4000 m. The temperatures can rise to as much as 20°C during the day. There are also vegetation zones similar to the Páramo at the heights of the African continent (e.g. Mount Kilimanjaro). Instead of the Espeletia and Puyas, the similarly adapted Dendrosenecio and Lobelia species can be found.
The phasmid fauna of the Páramo is amazingly rich in species. Usually, the species found there are very small (rarely more than 5 cm) or, like the highly specialized Monticomorpha species (Pseudophasmatinae), sturdily built with short limbs and coloured black. The dark colouring favours the admission of warmth through solar radiation.
Tierra Helada / Highland Desert>
Between the Páramo steppes and the snow line of the Andes, at an altitude of 4700 m, lies the almost vegetation-free zone of the Tierra Helada or Highland Desert. Highland deserts can be found in different parts of the world (Africa, the Himalayas & Andes), but phasmids in this zone are only known from the Andes. These are highly specialized species of the genus Monticomorpha, especially M. flavolimbata (Redtenbacher, 1906). This species is found at the Cotopaxi volcano in northern Ecuador at altitudes of up to 5000 m, where they live mostly on or under lava stones. These stones can store an amazing amount of heat due to the intense sunlight and are hand-warm even at air temperatures close to zero. The vegetation here is limited to isolated tufts of grasses, lichens and shallow cushion plants, which often have vivid red or yellow colour patterns.
Tropical Dry Forest / Dry Savannah
Tropical dry forests or seasonal forests occur in lowlands where rainfall is less abundant or where the dry season(s) extend over more than 2 months per year. In areas where rainy and dry seasons alternate, most trees lose their foliage at the beginning of the dry season and sprout again at the beginning of the rainy season. These forests, which are green only during a short rainy season, are therefore in some respects an ecological counterpart to the deciduous forests in Central Europe. Lianas and herbaceous epiphytes still occur in these dry forests, but the unfavourable water balance leads to low-growing trees, with more branching and smaller leaves. The biomass and biodiversity are considerably lower than in the rainforest. The diversity of species of phasmids is considerably lower in these areas. Nevertheless, a relatively high population density can be found for a few species if they are well-adapted to their environment.
If precipitation decreases even further and the dry seasons are prolonged, the forests grow even more sparsely. The trees are low-growing and the foliage is thicker and sparser. The undergrowth of such dry forests usually consists of grasses and herbaceous plants, which blend into tropical grasslands. This grassland is usually formed by tall, horst-forming grasses, with shrubs and trees growing individually or in groups. Such tropical or subtropical grasslands interspersed with isolated trees are generally referred to as savannahs. In eastern Africa or western Madagascar, where these tree clusters are mainly formed by acacias (Acacia spp., Mimosaceae), we speak of thorn savannahs. Savannahs can be found in Africa (Miombo forests) but also in South America (e.g. the so-called "Llanos" at Rio Orinoco and "Cerrado" in Brazil). The phasmid fauna of such areas is rather small, however, especially in the acacia forests of Africa or the west of Madagascar large phasmid forms have developed (e.g. Bactrododema species in Africa and Achrioptera species in Madagascar). Many of the highly adapted Gratidia species can be found in the grasslands of Africa.
Agricultural Land
Agriculturally used areas can be of a very different shape. In the immediate vicinity of cities, agricultural land is often used intensively, so that there are no or hardly any areas of primary or secondary forest. In rural areas far away from larger cities, land utilisation is less intensive, which is why smaller, isolated areas of secondary forest and various hedges or groups of trees often still exist. Agricultural land often borders on primary or secondary forest. The less primary and secondary forest there is in agar areas, the lower the number of phasmids occurring in such areas. In areas where agricultural land borders directly on primary forest areas, the phasmid collection is more productive and often surprisingly successful. This is probably due to the easily accessible and walkable forest edges with rich and varied vegetation near the ground. Even isolated hedges along roadsides and paths are quite productive for collecting, as long as there is still forest in the surrounding area.
Urban Areas
With their parks, hotel gardens or undeveloped plots of secondary forest, cities also offer potential opportunities for the presence of phasmids. In outlying areas with a more village character and more green areas or even areas with secondary forests, the presence of phasmids is even more likely. Since public parks and hotel gardens are intensively maintained and often treated with pesticides, it is rare to find a few phasmids there. Generally, it can be said that the population density increases with increasing distance from the city centre.
In the outskirts of Cayenne (French Guiana), for example, there are surprisingly high population densities with a relatively high species richness. Here, many phasmids can be found in the immediate vicinity of human dwellings and on various roadsides.